Приспособления, вырабатываемые жертвами для противодействия хищникам , способствуют выработке у хищников механизмов преодоления этих приспособлений. Длительное совместное существование хищников и жертв приводит к формированию системы взаимодействия, при которой обе группы устойчиво сохраняются на изучаемой территории. Нарушение такой системы часто приводит к отрицательным экологическим последствиям.
Негативное влияние нарушения коэволюционных связей наблюдается при интродукции видов. В частности, козы и кролики , интродуцированные в Австралии , не имеют на этом материке эффективных механизмов регуляции численности, что приводит к разрушению природных экосистем .
Математическая модель
Допустим, что на некоторой территории обитают два вида животных : кролики (питающиеся растениями) и лисы (питающиеся кроликами). Пусть число кроликов , число лис . Используя Модель Мальтуса с необходимыми поправками, учитывающими поедание кроликов лисами, приходим к следующей системе, носящей имя модели Вольтерры - Лотки :
\begin{cases} \dot x=(\alpha -c y)x;\\
\dot y=(-\beta+d x) y. \end{cases}
Поведение модели
Групповой образ жизни хищников и их жертв радикально меняет поведение модели, придает ей повышенную устойчивость.
Обоснование: при групповом образе жизни снижается частота случайных встреч хищников с потенциальными жертвами, что подтверждается наблюдениями за динамикой численности львов и антилоп гну в парке Серенгети .
История
Модель совместного существования двух биологических видов (популяций) типа «хищник - жертва» называется также моделью Вольтерры - Лотки.
См. также
Напишите отзыв о статье "Система «хищник - жертва»"
Примечания
Литература
- В. Вольтерра, Математическая теория борьбы за существование. Пер. с франц. О. Н. Бондаренко. Под ред и послесловием Ю. М. Свирежева. М.: Наука, 1976. 287 c. ISBN 5-93972-312-8
- А. Д. Базыкин, Математическая биофизика взаимодействующих популяций. М.: Наука, 1985. 181 с.
- А. Д. Базыкин, Ю. А. Кузнецов, А. И. Хибник, Портреты бифуркаций (Бифуркационные диаграммы- динамических систем на плоскости) /Серия «Новое в жизни, науке, технике. Математика, кибернетика» - М.: Знание, 1989. 48 с.
- П. В. Турчин,
Ссылки
Отрывок, характеризующий Система «хищник - жертва»
– Charmant, charmant, [Прелестно, прелестно,] – сказал князь Василий.– C"est la route de Varsovie peut etre, [Это варшавская дорога, может быть.] – громко и неожиданно сказал князь Ипполит. Все оглянулись на него, не понимая того, что он хотел сказать этим. Князь Ипполит тоже с веселым удивлением оглядывался вокруг себя. Он так же, как и другие, не понимал того, что значили сказанные им слова. Он во время своей дипломатической карьеры не раз замечал, что таким образом сказанные вдруг слова оказывались очень остроумны, и он на всякий случай сказал эти слова, первые пришедшие ему на язык. «Может, выйдет очень хорошо, – думал он, – а ежели не выйдет, они там сумеют это устроить». Действительно, в то время как воцарилось неловкое молчание, вошло то недостаточно патриотическое лицо, которого ждала для обращения Анна Павловна, и она, улыбаясь и погрозив пальцем Ипполиту, пригласила князя Василия к столу, и, поднося ему две свечи и рукопись, попросила его начать. Все замолкло.
– Всемилостивейший государь император! – строго провозгласил князь Василий и оглянул публику, как будто спрашивая, не имеет ли кто сказать что нибудь против этого. Но никто ничего не сказал. – «Первопрестольный град Москва, Новый Иерусалим, приемлет Христа своего, – вдруг ударил он на слове своего, – яко мать во объятия усердных сынов своих, и сквозь возникающую мглу, провидя блистательную славу твоея державы, поет в восторге: «Осанна, благословен грядый!» – Князь Василий плачущим голосом произнес эти последние слова.
Билибин рассматривал внимательно свои ногти, и многие, видимо, робели, как бы спрашивая, в чем же они виноваты? Анна Павловна шепотом повторяла уже вперед, как старушка молитву причастия: «Пусть дерзкий и наглый Голиаф…» – прошептала она.
Князь Василий продолжал:
– «Пусть дерзкий и наглый Голиаф от пределов Франции обносит на краях России смертоносные ужасы; кроткая вера, сия праща российского Давида, сразит внезапно главу кровожаждущей его гордыни. Се образ преподобного Сергия, древнего ревнителя о благе нашего отечества, приносится вашему императорскому величеству. Болезную, что слабеющие мои силы препятствуют мне насладиться любезнейшим вашим лицезрением. Теплые воссылаю к небесам молитвы, да всесильный возвеличит род правых и исполнит во благих желания вашего величества».
– Quelle force! Quel style! [Какая сила! Какой слог!] – послышались похвалы чтецу и сочинителю. Воодушевленные этой речью, гости Анны Павловны долго еще говорили о положении отечества и делали различные предположения об исходе сражения, которое на днях должно было быть дано.
– Vous verrez, [Вы увидите.] – сказала Анна Павловна, – что завтра, в день рождения государя, мы получим известие. У меня есть хорошее предчувствие.
Предчувствие Анны Павловны действительно оправдалось. На другой день, во время молебствия во дворце по случаю дня рождения государя, князь Волконский был вызван из церкви и получил конверт от князя Кутузова. Это было донесение Кутузова, писанное в день сражения из Татариновой. Кутузов писал, что русские не отступили ни на шаг, что французы потеряли гораздо более нашего, что он доносит второпях с поля сражения, не успев еще собрать последних сведений. Стало быть, это была победа. И тотчас же, не выходя из храма, была воздана творцу благодарность за его помощь и за победу.
Предчувствие Анны Павловны оправдалось, и в городе все утро царствовало радостно праздничное настроение духа. Все признавали победу совершенною, и некоторые уже говорили о пленении самого Наполеона, о низложении его и избрании новой главы для Франции.
Вдали от дела и среди условий придворной жизни весьма трудно, чтобы события отражались во всей их полноте и силе. Невольно события общие группируются около одного какого нибудь частного случая. Так теперь главная радость придворных заключалась столько же в том, что мы победили, сколько и в том, что известие об этой победе пришлось именно в день рождения государя. Это было как удавшийся сюрприз. В известии Кутузова сказано было тоже о потерях русских, и в числе их названы Тучков, Багратион, Кутайсов. Тоже и печальная сторона события невольно в здешнем, петербургском мире сгруппировалась около одного события – смерти Кутайсова. Его все знали, государь любил его, он был молод и интересен. В этот день все встречались с словами:
– Как удивительно случилось. В самый молебен. А какая потеря Кутайсов! Ах, как жаль!
– Что я вам говорил про Кутузова? – говорил теперь князь Василий с гордостью пророка. – Я говорил всегда, что он один способен победить Наполеона.
Но на другой день не получалось известия из армии, и общий голос стал тревожен. Придворные страдали за страдания неизвестности, в которой находился государь.
– Каково положение государя! – говорили придворные и уже не превозносили, как третьего дня, а теперь осуждали Кутузова, бывшего причиной беспокойства государя. Князь Василий в этот день уже не хвастался более своим protege Кутузовым, а хранил молчание, когда речь заходила о главнокомандующем. Кроме того, к вечеру этого дня как будто все соединилось для того, чтобы повергнуть в тревогу и беспокойство петербургских жителей: присоединилась еще одна страшная новость. Графиня Елена Безухова скоропостижно умерла от этой страшной болезни, которую так приятно было выговаривать. Официально в больших обществах все говорили, что графиня Безухова умерла от страшного припадка angine pectorale [грудной ангины], но в интимных кружках рассказывали подробности о том, как le medecin intime de la Reine d"Espagne [лейб медик королевы испанской] предписал Элен небольшие дозы какого то лекарства для произведения известного действия; но как Элен, мучимая тем, что старый граф подозревал ее, и тем, что муж, которому она писала (этот несчастный развратный Пьер), не отвечал ей, вдруг приняла огромную дозу выписанного ей лекарства и умерла в мучениях, прежде чем могли подать помощь. Рассказывали, что князь Василий и старый граф взялись было за итальянца; но итальянец показал такие записки от несчастной покойницы, что его тотчас же отпустили.
Математическое моделирование биологических процессов началось с создания первых простейших моделей экологической системы.
Допустим, в некотором замкнутом районе живут рыси и зайцы. Рыси питаются только зайцами, а зайцы – растительной пищей, имеющейся в неограниченном количестве. Необходимо найти макроскопические характеристики, описывающие популяции. Такими характеристиками являются число особей в популяциях.
Простейшая модель взаимоотношений популяций хищника и жертвы, основанная на логистическом уравнении роста, названа (как и модель межвидовой конкуренции) по имени ее создателей - Лотки и Вольтерра. Эта модель крайне упрощает исследуемую ситуацию, но все же полезна в качестве отправной точки в анализе системы хищник-жертва.
Предположим, что (1) популяция жертвы существует в идеальной (независимой от плотности) среде, где ее рост может ограничивать только наличие хищника, (2) столь же идеальна среда, в которой существует хищник, рост популяции которого ограничивает лишь обилие жертв, (3) обе популяции размножаются непрерывно согласно экспоненциальному уравнению роста, (4) скорость поедания жертв хищниками пропорциональна частоте встреч между ними, которая, в свою очередь, является функцией плотности популяций. Эти допущения и лежат в основе модели Лотки - Вольтерра.
Пусть в отсутствие хищников популяция жертвы растет экспоненциально:
dN/dt =r 1 N 1
где N -численность, а r, - удельная мгновенная скорость роста популяции жертвы. Если же хищники присутствуют, то они уничтожают особей жертвы со скоростью, которая определяется, во-первых, частотой встреч хищников и жертв, возрастающей по мере увеличения их численностей, и, во-вторых, эффективностью, с которой хищник обнаруживает и ловит свою жертву при встрече. Число жертв, встреченных и съеденных одним хищником N с, пропорционально эффективности охоты, которую мы выразим через коэффициент С 1; численности (плотности) жертвы N и времени, затраченному на поиски Т:
N C =C 1 NT (1)
Из этого выражения легко определить удельную скорость потребления жертв хищником (т.е. число жертв, поедаемых одной особью хищника в единицу времени), которую часто называют также функциональным ответом хищника на плотность популяции жертвы:
В рассматриваемой модели С 1 является константой. Это означает, что число жертв, изъятых хищниками из популяции, линейно возрастает с увеличением ее плотности (так называемый функциональный ответ типа 1). Ясно, что общая скорость поедания жертв всеми особями хищника составит:
![]() (3)
(3)
где Р - численность популяции хищника. Теперь мы можем записать уравнение роста популяции жертвы следующим образом:
При отсутствии жертвы особи хищника голодают и гибнут. Предположим также, что в этом случае численность популяции хищника будет уменьшаться экспоненциально согласно уравнению:
![]() (5)
(5)
где r 2 - удельная мгновенная смертность в популяции хищника.
Если жертвы присутствуют, то те особи хищника, которые смогут их найти и съесть, будут размножаться. Рождаемость в популяции хищника в данной модели зависит только от двух обстоятельств: скорости потребления жертв хищником и эффективности, с которой поглощенная пища перерабатывается хищником в его потомство. Если мы выразим эту эффективность через коэффициент s, то рождаемость составит:
![]()
Поскольку С 1 и s - константы, их произведение - это также константа, которую мы обозначим как С 2 . Тогда скорость роста популяции хищника будет определяться балансом рождаемости и смертности в соответствии с уравнением:
![]() (6)
(6)
Уравнения 4 и 6 вместе образуют модель Лотки-Вольтерра.
Свойства этой модели мы можем исследовать точно так же, как и в случае конкуренции, т.е. построив фазовую диаграмму, на которой численность жертвы отложена по оси ординат, а хищника - по оси абсцисс, и проведя на ней изоклины-линии, соответствующие постоянной численности популяций. С помощью таких изоклин определяют поведение взаимодействующих популяций хищника и жертвы.
Для популяции жертвы: при откуда
Таким образом, поскольку r, и С 1 , - константы, изоклиной для жертвы будет линия, на которой численность хищника (Р) является постоянной, т.е. параллельная оси абсцисс и пересекающая ось ординат в точке Р =r 1 / С 1 . Выше этой линии численность жертвы будет уменьшаться, а ниже- возрастать.
Для популяции хищника:
при откуда
Поскольку r 2 и С 2 - константы, изоклиной для хищника будет линия, на которой численность жертвы (N) является постоянной, т.е. перпендикулярная оси ординат и пересекающая ось абсцисс в точке N = r 2 /С 2 . Слева от нее численность хищника будет уменьшаться, а справа - возрастать.
Если мы рассмотрим эти две изоклины вместе, то легко заметим, что взаимодействие популяций хищника и жертвы имеет циклический характер, поскольку их численности претерпевают неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищничества на популяцию жертвы и тем самым к снижению ее численности. Это снижение, в свою очередь, ведет к нехватке пищи у хищников и падению их численности, которое вызывает ослабление пресса хищничества и увеличению численности жертвы, что снова приводит к росту популяции жертвы и т.д.
Для данной модели характерна так называемая "нейтральная стабильность", которая означает, что популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новый цикл колебаний с иными параметрами. Для того, чтобы циклы стали стабильными, популяции должны после внешнего воздействия стремиться вернуться к первоначальному циклу. Такие циклы, в отличие от нейтрально стабильных колебаний в модели Лотки-Вольтерра, принято называть устойчивыми предельными циклами.
Модель Лотки-Вольтерра, тем не менее, полезна тем, что позволяет продемонстрировать основную тенденцию в отношениях хищник-жертва возникновение циклических сопряженных колебаний численности их популяций.
к договору от ___.___,20___ г. об оказании платных образовательных услуг
Министерство образования и науки Российской Федерации
Лысьвенский филиал
Пермского Государственного технического университета
Кафедра ЕН
Курсовая работа
по дисциплине «Моделирование систем»
тема: Система хищник-жертва
Выполнил:
Студент гр. БИВТ-06
------------------
Проверил преподаватель:
Шестаков А. П.
Лысьва, 2010 г.
Реферат
Хищничество - трофические отношения между организмами, при которых один из них (хищник) атакует другого (жертву) и питается частями его тела, то есть обычно присутствует акт умерщвления жертвы. Хищничество противопоставляется поеданию трупов (некрофагии) и органических продуктов их разложения (детритофагии).
Довольно популярно также другое определение хищничества, предлагающее хищниками называть лишь организмы, поедающие животных, в отличие от растительноядных, поедающих растения.
Кроме многоклеточных животных, в роли хищников могут выступать протисты, грибы и высшие растения.
Размер популяции хищников влияет на размер популяции их жертв и наоборот, динамика популяций описывается математической моделью Лотки-Вольтерра, однако данная модель является высокой степенью абстракции, и не описывает реальные взаимосвязи между хищником и жертвой, и может рассматриваться только как первая степень приближения математической абстракции.
В процессе совместной эволюции хищники и жертвы приспосабливаются друг к другу. У хищников появляются и развиваются средства обнаружения и атаки, а у жертв - средства скрытности и защиты. Поэтому наибольший вред жертвам могут нанести новые для них хищники, с которыми те не вступали ещё в «гонку вооружений».
Хищники могут специализироваться на одном–нескольких видах для добычи, это делает их в среднем более успешными в охоте, но повышает зависимость от данных видов.
Система хищник-жертва.
Взаимодействие хищник-жертва - основной тип вертикальных взаимоотношений организмов, при котором по пищевым цепям передаются вещество и энергия.
Равновесность В. х. - ж. наиболее легко достигается в том случае, если в пищевой цепи имеется не менее трех звеньев (например, трава - полёвка - лисица). При этом плотность популяции фитофага регулируется взаимоотношениями как с нижним, так и с верхним звеном пищевой цепи.
В зависимости от характера жертв и типа хищника (истинный, пастбищник) возможна разная зависимость динамики их популяций. При этом картина осложняется тем, что хищники очень редко бывают монофагами (т. е. питающимися одним видом жертвы). Чаще всего, когда истощается популяция одного вида жертвы и ее добывание требует слишком больших затрат сил, хищники переключаются на другие виды жертв. Кроме того, одну популяцию жертв может эксплуатировать несколько видов хищников.
По этой причине часто описываемый в экологической литературе эффект пульсирования численности популяции жертвы, за которым с определенным запаздыванием пульсирует численность популяции хищника, в природе встречается крайне редко.
Равновесие между хищниками и жертвами у животных поддерживается специальными механизмами, исключающими полное истребление жертв. Так, жертвы могут:
- убегать от хищника (в этом случае в результате соревнования повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей);
- приобретать защитную окраску (<притворяться> листьями или сучками) или, напротив, яркий (например, красный) цвет, предупреждающий хищника о горьком вкусе;
- прятаться в укрытия;
- переходить к мерам активной обороны (рогатые травоядные, колючие рыбы), часто совместной (птицы-жертвы коллективно отгоняют коршуна, самцы оленей и сайгаков занимают <круговую оборону> от волков и т. д.).
Хищники могут поедать травоядных животных, и также слабых хищников. Хищники обладают широким спектром питания, легко переключаются с одной добычи на другую, более доступную. Хищники часто нападают на слабые жертвы. Поддерживается экологическое равновесие между популяциями жертва-хищник.[ ...]
Если равновесие неустойчиво (предельных циклов нет) или внешний цикл неустойчив, то численности обоих видов, испытывая сильные колебания, уходят из окрестности равновесия. Причем быстрое вырождение (в первой ситуации) наступает при низкой адаптации хищника, т.е. при его высокой смертности (по сравнению со скоростью размножения жертвы). Это означает, что слабый во всех отношениях хищник не способствует стабилизации системы и сам вымирает.[ ...]
Пресс хищников особенно силен, когда в коэволюции хищник - жертва равновесие смещается в сторону хищника и ареал жертвы сужается. Конкурентная борьба тесно связана с нехваткой пищевых ресурсов, она может быть и прямой борьбой, например, хищников за пространство как ресурс, но чаще всего это просто вытеснение вида, которому на данной территории пищи не хватает, видом, которому этого же количества пищи вполне достаточно. Это уже межвидовая конкуренция.[ ...]
 |
И окончательно в системе ’’хищник - жертва”, описываемой моделью (2.7), возникновение диффузионной неустойчивости (при локальной устойчивости равновесия) возможно лишь в том случае, когда естественная смертность хищника возрастает с ростом его численности быстрее, чем линейная функция, и трофическая функция отличается от вольтерровской либо, когда популяция жертвы - это популяция типа Олли.[ ...]
Теоретически в моделях «один хищник - две жертвы» эквивалентное выедание (отсутствие предпочтения того или иного вида жертвы) может повлиять на конкурентное сосуществование видов-жертв лишь в тех местах, где уже существует потенциально устойчивое равновесие . Разнообразие может возрасти только в таких условиях, когда у видов с меньшей конкурентоспособностью скорость роста популяции выше, чем у доминантов. Это позволяет понять ситуацию, когда равномерное выедание ведет к увеличению видового разнообразия растений там, где большее число видов, прошедших отбор на быстрое размножение, сосуществует с видами, эволюция которых направлена на повышение конкурентоспособности .[ ...]
Точно так же выбор жертвы, зависящий от ее плотности, может привести к устойчивому равновесию в теоретических моделях двух конкурирующих видов жертвы, где прежде никакого равновесия не существовало . Для этого хищник должен был бы обладать способностью к функциональным и численным реакциям на изменения плотности жертвы; возможно, однако, что переключение (непропорционально частые нападения на наиболее обильную жертву) будет иметь при этом более важное значение. В самом деле, установлено, что переключение оказывает стабилизирующее влияние в системах «один хищник - п жертв» и представляет собой единственный механизм, способный стабилизировать взаимодействия в тех случаях, когда ниши жертв полностью перекрываются . Такую роль могут играть неспециализированные хищники. Предпочтение более специализированными хищниками доминантного конкурента действует таким же образом, как переключение хищника, и может стабилизировать теоретические взаимодействия в моделях, в которых прежде не существовало равновесия между видами жертвы, при условии что их ниши в какой-то степени разделены .[ ...]
Также- не стабилизирует сообщество и хищник ’’сильный во всех отношениях”, т.е. хорошо адаптированный к данной жертве и с низкой относительной смертностью. При этом система обладает неустойчивым предельным циклом и, несмотря на устойчивость положения равновесия, вырождается в случайной среде (хищник выедает жертву и вследствие этого гибнет). Такая ситуация соответствует медленному вырождению.[ ...]
Таким образом, при хорошей адаптации хищника в окрестности устойчивого равновесия могут возникать неустойчивый и устойчивый циклы, т.е. в зависимости от начальных условий система ’’хищник-жертва” либо стремится к равновесию, либо, колеблясь, уходит от него, либо в окрестности равновесия устанавливаются устойчивые колебания численностей обоих видов.[ ...]
Организмы, которых относят к хищникам, питаются другими организмами, уничтожая свою жертву. Таким образом, среди живых организмов следует выделять еще одну классификационную систему, а именно «хищники» и «жертвы». Отношения между такими организмами складывались на протяжении всей эволюции жизни на нашей планете. Организмы-хищники выступают в роли природных регуляторов численности организмов-жертв. Увеличение численности «хищников» приводит к уменьшению численности «жертв», это, в свою очередь, снижает запасы пищи («жертв») для «хищников», что в целом диктует снижение численности «жертв» и т. д. Таким образом, в биоценозе постоянно происходят колебания численности хищников и жертв, в целом же устанавливается определенное равновесие на какой-то период времени в пределах достаточно устойчивых условий среды.[ ...]
Это в конечном итоге приходит к экологическому равновесию между популяциями хищника и жертвы.[ ...]
Для трофической функции третьего типа состояние равновесия будет устойчивым, если где N - точка перегиба функции (см. рис. 2, в). Это следует из того, что на интервале трофическая функция вогнута и, следовательно, относительная доля потребления хищником жертвы возрастает.[ ...]
Пусть Гг = -Г, т.е. имеет место сообщества типа ’’хищник - жертва”. В этом случае первое слагаемое в выражении (7.4) равно нулю и для выполнения условия устойчивости по вероятности равновесного состояния N требуется, чтобы и второе слагаемое не было положительным.[ ...]
Таким образом, для рассмотренного сообщества типа хищник - жертва можно сделать вывод об асимптотической устойчивости в целом положительного положения равновесия, т. е. для любых начальных данных 1Ч(0)>0 эволюция происходит таким образом, что N(7) - ■ К при условии, что N >0.[ ...]
Так, в однородной среде, не имеющей укрытий для размножения, хищник рано или поздно уничтожает популяцию жертвы и после этого вымирает сам. Волны жизни” (изменений численности хищника и жертвы) следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника, так и жертвы остается примерно на одном уровне. Длительность периода зависит от скоростей роста обоих видов и от исходных параметров. Для популяции жертвы влияние хищника положительно, так как ее чрезмерное размножение привело бы к краху ее численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.[ ...]
Другие модификации могут быть следствием особенностей поведения хищника. Число особей жертвы, которое хищник в состоянии потребить в данное время, имеет свой предел. Эффект насыщения хищника при приближении к этому рубежу показан в табл. 2-4, В. Взаимодействия, описываемые уравнениями 5 и 6, могут иметь устойчивые точки равновесия или обнаруживать циклические колебания. Однако такие циклы отличаются от тех, какие отражены в уравнениях Лотки - Вольтерры 1 и 2. Циклы, передаваемые уравнениями 5 и 6, могут иметь постоянную амплитуду и средние плотности, пока среда постоянна; после того как произошло нарушение, они могут вернуться к прежним амплитудам и средним плотностям. Такие циклы, которые восстанавливаются после нарушений, называются устойчивыми предельными циклами. Взаимодействие зайца и рыси можно считать устойчивым предельным циклом, но это не цикл Лотки - Вольтерры.[ ...]
Рассмотрим возникновение диффузионной неустойчивости в системе ’’хищник -жертва”, но сначапа выпишем условия, обеспечивающие возникновение диффузионной неустойчивости в системе (1.1) при п = 2. Ясно, что равновесие (N , Щ) локально (т.е.[ ...]
Перейдем к интерпретации случаев, связанных с длительным сосуществованием хищника и жертвы. Понятно, что в отсутствие предельных циклов устойчивому равновесию будут соответствовать в случайной среде флуктуации численности, причем их амплитуда будет пропорциональна дисперсии возмущений. Такое явление будет происходить, если хищник имеет высокую относительную смертность и в то же время высокую степень приспособленности к данной жертве.[ ...]
Рассмотрим теперь, как меняется динамика системы с ростом приспособленности хищника, т.е. с убыванием Ъ от 1 до 0. Если приспособленность достаточно низкая, то предельные циклы отсутствуют, а равновесие является неустойчивым. С ростом приспособленности в окрестности этого равновесия возможно появление устойчивого цикла и далее внешнего неустойчивого. В зависимости от начальных условий (соотношения биомассы хищника и жертвы) система может либо терять устойчивость, т.е. уходить из окрестности равновесия, либо в ней будут со временем устанавливаться устойчивые колебания. Дальнейший рост приспособленности делает невозможным колебательный характер поведения системы. Однако при Ъ [ ...]
Пример отрицательной (стабилизирующей) обратной связи - взаимоотношение между хищником и жертвой или функционирование карбонатной системы океана (раствор СОг в воде: СО2 + Н2О -> Н2СОз). Обычно количество углекислоты, растворенное в воде океана, находится в парциальном равновесии с концентрацией углекислого газа в атмосфере. Локальные увеличения углекислоты в атмосфере после извержения вулканов приводят к интенсификации фотосинтеза и поглощению ее карбонатной системой океана. При снижении уровня углекислого газа в атмосфере карбонатная система океана высвобождает СОг в атмосферу. Поэтому концентрация углекислого газа в атмосфере достаточно стабильна.[ ...]
[ ...]
Как отмечает Р.Риклефс (1979), существуют факторы, способствующие стабилизации взаимоотношений в системе “хищник-жертва”: неэффективность хищника, наличие у хищника альтернативных пищевых ресурсов, уменьшение запаздывания в реакции хищника, а также экологические ограничения, налагаемые внешней средой на ту или иную популяцию. Взаимодействия между популяциями хищника и жертвы весьма разнообразны и сложны. Так, если хищники достаточно эффективны, они могут регулировать плотность популяции жертвы, удерживая ее на уровне ниже емкости среды. Через влияние, оказываемое ими на популяции жертвы, хищники воздействуют на эволюцию различных признаков жертвы, что приводит в конечном итоге к экологическому равновесию между популяциями хищника и жертвы.[ ...]
Если выполняется одно из условий: 0 1/2. Если же 6 > 1 (кА [ ...]
Устойчивость биоты и окружающей среды зависит только от взаимодействия растений - автотрофов и растительноядных гетеротрофных организмов. Хищники любых размеров не способны нарушить экологическое равновесие сообщества, так как в естественных условиях они не могут увеличить свою численность при постоянной численности жертв. Хищники не только должны быть сами передвигающимися, но и могут питаться только передвигающимися животными.[ ...]
Никакие другие рыбы не распространены так широко, как щуки. В немногих местах товли в стоячих или проточных водоемах нет давления со стороны щук для сохранения равновесия между жертвой и хищником Только современные искусственные водоемы, в которых щуки являются рыбами нежелательными из-за разведения там других рыб, не заселяются ими целенаправленно. В мире щуки исключительно хорошо представлены. Их ловят по всему северном) полушарию от Соединенных Штатов и Канады в Северной Америке, через Европу до северной части Азии.[ ...]
Еще одна возможность устойчивого сосуществования возникает здесь же, в узком диапазоне сравнительно высокой адаптации. При переходе к неустойчивому режиму с очень ’’хорошим” хищником может возникнуть устойчивый внешний предельный цикл, при котором диссипация биомассы уравновешивается ее притоком в систему (высокой продуктивностью жертвы). Тогда возникает любопытная ситуация, когда наиболее вероятными являются два характерных значения амплитуды случайных колебаний. Одни происходят вблизи равновесия, другие - вблизи предельного цикла, причем возможны более или менее частые переходы между этими режимами.[ ...]
Гипотетические популяции, которые ведут себя в соответствии с векторами на рис. 10.11 А, изображены на рис. 10.11,-Б с помощью графика, показывающего динамику соотношения численностей хищника и жертвы и на рис. 10.11,5 в виде графика динамики численности хищника и жертвы во времени. В популяции жертвы, по мере того как она переходит от равновесия прн низкой плотности к равновесию при высокой и возвращается назад, происходит «вспышка» численности. И эта вспышка численности не является следствием столь же выраженного изменения в окружающей среде. Напротив, это изменение численности порождено самим воздействием (при небольшом уровне «шума» в среде) и оно, в частности, отражает существование нескольких равновесных состояний. Сходные рассуждения можно использовать для объяснения более сложных случаев динамики численности в природных популяциях.[ ...]
Важнейшим свойством экосистемы является ее устойчивость, сбалансированность обмена и происходящих в ней процессов. Способность популяций или экосистемы поддерживать устойчивое динамическое равновесие в изменяющихся условиях среды называется гомеостазом (homoios - тот же, подобный; stasis - состояние). В основе гомеостаза лежит принцип обратной связи. Для поддержания равновесия в природе не требуется внешнего управления. Пример гомеостаза - субсистема «хищник-жертва», в которой регулируются плотность популяций хищника и жертвы.[ ...]
Природная экосистема (биогеоценоз) устойчиво функционирует при постоянном взаимодействии ее элементов, круговороте веществ, передаче химической, энергетической, генетической и другой энергии и информации по цепям-каналам. Согласно принципу равновесия любая естественная система с проходящим через нее потоком энергии и информации имеет тенденцию к развитию устойчивого состояния. При этом устойчивость экосистем обеспечивается автоматически за счет механизма обратной связи. Обратная связь заключается в использовании получаемых данных от управляемых компонентов экосистем для внесения корректив управляющими компонентами в процесс. Рассмотренные выше взаимоотношения «хищник» - «жертва» в данном контексте можно описать несколько подробнее; так, в водной экосистеме хищные рыбы (щука в пруду) поедают другие виды рыб-жертвы (карась); если численность карася будет увеличиваться -это пример положительной обратной связи; щука, питаясь карасем, снижает его численность-это пример отрицательной обратной связи; при росте числа хищников снижается число жертв, и хищник, испытывая недостаток пищи, также снижает рост своей популяции; в конце концов в рассматриваемом пруду устанавливается динамическое равновесие в численности и щуки, и карася. Постоянно поддерживается равновесие, которое исключало бы исчезновение любого звена трофической цепи (рис. 64).[ ...]
Переходим к наиболее важному обобщению, а именно что отрицательные взаимодействия со временем становятся менее заметными, если экосистема достаточно стабильна и ее пространственная структура обеспечивает возможность взаимного приспособления популяций. В модельных системах типа хищник- жертва, описываемых уравнением Лотки-Вольтерры, если в уравнение не введены дополнительные члены, характеризующие действие факторов самоограничения численности, то колебания происходят непрерывно и не затухают (см. Левонтин, 1969). Пиментел (1968; см. также Пиментел и Стоун, 1968) экспериментально показал, что такие дополнительные члены могут отражать взаимные адаптации или генетическую обратную связь. Когда же новые культуры создавали из особей, ранее на протяжении двух лет совместно существовавших в культуре, где их численность подвергалась значительным колебаниям, оказалось, что у них выработался экологический гомеостаз, при котором каждая из популяций была «подавлена» другой в такой степени, что оказалось возможным их сосуществование при более стабильном равновесии.
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
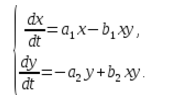
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).